Bottom
The fungal pathogen Candida albicans is commonly seen in immunosuppressed patients, and resistance to one of the most widely used antifungals, fluconazole (FLC), can evolve rapidly. In recent years it has become clear that the plasticity of the Candida albicans genome contributes to drug resistance through loss of heterozygosity (LOH) in resistance genes and macroscopic chromosomal rearrangements that amplify gene copy number. associated with resistance. This study addresses the role of the homologous recombination factors Rad54 and Rdh54 in cell growth, DNA damage and resistance to FLC in Candida albicans.
Materials And Methods
- Strains and growth conditions.
Candida albicans recombinants wild-type strain SC5314 was used to construct all mutants created for this study. Deletion and replacement of Candida albicans RAD54 and Candida albicans RDH54 was performed using the nourseothricin resistance marker SAT1 (generously provided by Dr Joachim Morchauser) to create homozygous null mutants Candida albicans rdh54Δ/rdh54Δ, Candida albicans rad54Δ/rad54Δ and the reconstructed strain Candida albicans rad54Δ/RAD54 (+).
The rad54Δ/RAD54 (+) reconstructed strain was prepared from one of the rad54Δ/rad54Δ strains. For routine growth, strains were maintained at 30°C in YPD (10 g Difco yeast extract, 20 g Bacto peptone and 20 g dextrose per litre) with or without 200 µg/ml nourseothricin. Spider medium was used for agar invasion tests, with a final pH of 7.2 (10 g nutrient broth, 10 g mannitol, 2 g K2PO4 and 25 g agar per litre).
- Plasmid Construction
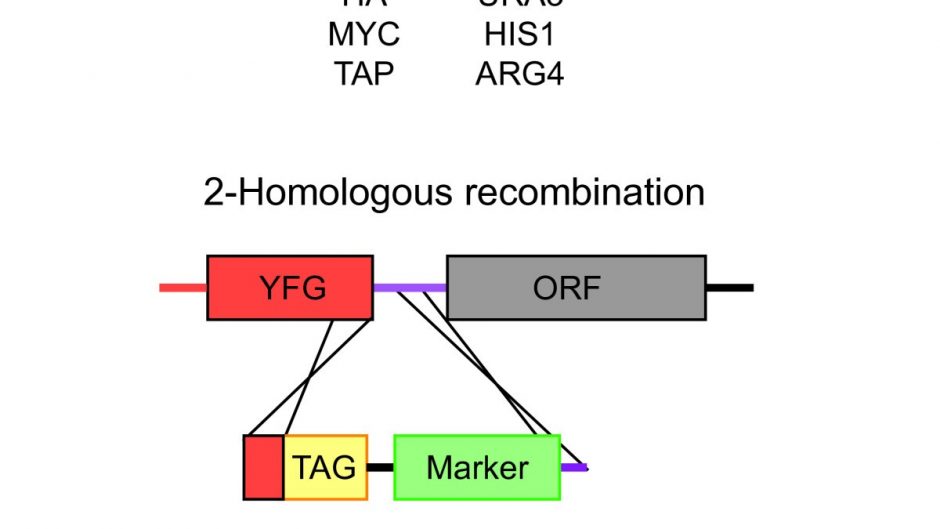
To create null mutants of Candida albicans RAD54 and Candida albicans RDH54, approximately 500 base pairs (bp) of sequence upstream and downstream of these ORFs were cloned on either side of the SAT1-FLP cassette into the pFS2A vector. Fragments were designed such that the entire coding sequence from ATG to stop codon was replaced by the SAT1 cassette. For both genes, the upstream fragment was cloned using the restriction enzymes ApaI and XhoI and the downstream fragment was cloned using NotI and SacII. To create the Candida albicans RAD54 reconstruction vector, the entire coding region, including the promoter and terminator sequence, was cloned into the ApaI-XhoI site in the Candida albicans RAD54 deletion vector.
- Yeast transformation and screening
SC5314 was transformed with linearized Candida albicans RAD54 or Candida albicans RDH54 deletion vectors (linearized with ApaI and SacII) using the standard lithium acetate method with the following modifications. Heat shock was performed at 42°C overnight, and cells were resuspended in YPD and allowed to grow for 4 hours at 30°C before seeding on YPD containing 200 µg/mL cloNAT (Werner BioAgents, Jenna, Germany). Recycling of the SAT1 marker was performed by culturing cells overnight in non-selective media (YPD) and plating on YPD containing 25 µg/ml nourseothricin.
Small colonies that had the marker removed were screened by PCR and used in a successive round of transformation. These transformants were then selected by PCR for the homozygous deletion of Candida albicans RAD54 and Candida albicans RDH54. To create the Candida albicans reconstruction strain RAD54, recycling of the SAT1 marker was performed again and the reconstruction plasmid was introduced into the native locus by another round of transformation.
- Determination of growth rate
Overnight YPD cultures of three independent colonies were used to inoculate 3 ml of YPD at an OD600 of 0.05. Cultures were grown at 30°C with shaking. OD measurements were taken every hour for 9 hours to generate growth curves. Doubling times for each strain were calculated using time points within the log phase of growth. This test was repeated three times, the mean and standard deviations for each strain are shown.
- Colony morphology and microscopic analysis
For evaluation of colony morphology, cells were grown in YPD for 2 days at 30°C and single colonies were photographed. For agar colony invasion, strains were plated on Spider agar plates (1% nutrient broth, 1% mannitol, 0.2% K2HPO4, and 20 g Bacto agar per litre) and incubated at 37°C. C for seven days and images were taken. For cell morphology, cells were grown in YPD to early log phase from overnight YPD cultures. Samples were taken, washed and resuspended in PBS buffer and sonicated for 5 seconds at 30% amplitude in a Fisher Scientific 150T Series sonic dismemberer (Fisher Scientific, USA).
Light microscopy was used to quantify the number of individual non-budding cells, budding cells, and cells with abnormal or pseudohyphal-like morphology. To assess nuclear integrity, cells were grown to the early log phase and stained with DAPI according to a previously published protocol. Overnight cultures were diluted to an OD600 of 0.05 in 5 mL YPD and grown for 4 hours at 30°C. Samples were centrifuged, washed in 1 ml 1X PBS and fixed overnight at 4°C in 1 ml 70% ethanol. Fixed cells were washed and treated for 2 hours in 55 mM HCl with 5 mg/ml pepsin at 37 °C, then washed and resuspended in 1 ml 1X PBS containing 2.5 μg/ml DAPI (Sigma- Aldrich, St. Louis, MO, USA).
- DNA damage and sensitivities to antifungal drugs
To test the sensitivity of the strains in this study to various agents, the agar spot dilution method was used. Overnight YPD cultures were diluted to an OD 600 of 1.0 and ten-fold serial dilutions were made to 10-6. Volumes of 2 μL of each dilution were plated onto YPD plates and YPD plates containing FLC, MMS, or menadione (Sigma-Aldrich, St. Louis, MO, USA) at the indicated concentrations. Plates were incubated for 48 hours at 30°C and images were taken.
An E-test analysis for common antifungals was performed, using overnight cultures diluted to an OD600 of 0.05 to spread a lawn on CAS plates (9.0 g casitone, 5.0 g yeast extract, 0. 54 g of KH2PO4, 3.34 g of K2HPO4, 20.0 g of dextrose and 20.0 g of agar per litre). Test strips E were plated on plates, which were incubated for 48 hours at 30°C. Two independent nulls of the RAD54 gene were tested. The MIC was read as the point where the zone of inhibition intersected test strip E.
Results
The data presented here support a role for homologous recombination in cell growth and sensitivity to DNA damage, as Candida albicans rad54Δ/rad54Δ mutants were hypersensitive to MMS and menadione, and had aberrant cellular and nuclear morphology. The Candida albicans rad54Δ/rad54Δ mutant was defective in the invasion of Spider agar, presumably due to altered cell morphology. In contrast, mutation of the related gene RDH54 did not contribute significantly to DNA damage resistance or cell growth, and deletion of Candida albicans RAD54 or Candida albicans RDH54 did not alter susceptibility to FLC.
Conclusions
Together, these results support a role for homologous recombination in genome stability under non-damaging conditions. Nuclear morphology defects in rad54Δ/rad54Δ mutants show that Rad54 plays an essential role during mitotic growth and that, in its absence, cells arrest in G2. The viability of the rad54Δ/rad54Δ single mutant and the inability to construct the rad54Δ/rad54Δ rdh54Δ/rdh54Δ double mutant suggests that Rdh54 may partially compensate for Rad54 during mitotic growth.